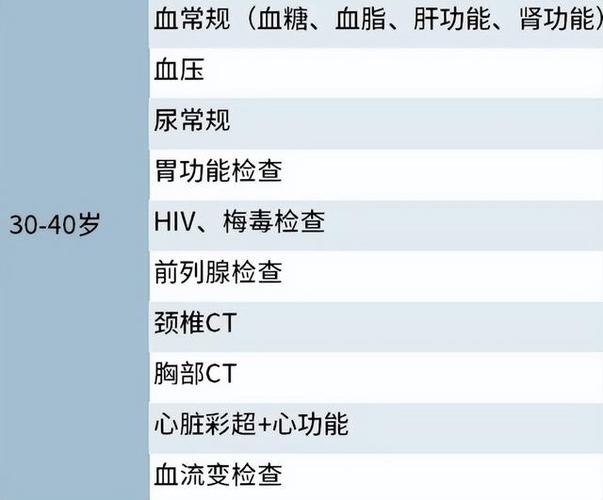
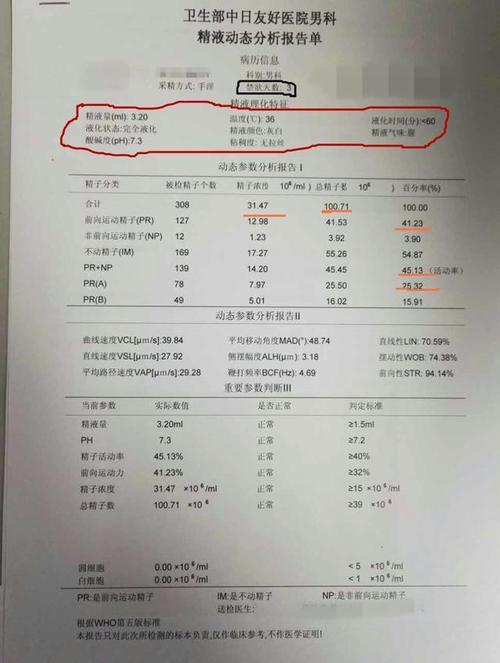
脂肪干细胞胶是什么
近年来,以干细胞为核心的再生医疗项目备受关注,从抗衰老、皮肤毛发再生、性功能改善,再到骨骼再生、免疫力提升等,其以促进组织再生或自我修复的治疗模式为人类带来了无限可能。
目前在日本干细胞治疗中,最受欢迎的、效果最显着的、风险最低的就是干细胞上清液疗法。

什么是脂肪干细胞上清液疗法?
脂肪干细胞上清液疗法,是将脂肪干细胞分泌出来的复数的成长因子以及胶原蛋白、玻尿酸等数百种蛋白质用于实际医疗中的治疗方法。
最大限度的使用人类自身的再生能力,来调和自身的成长因子,是安全有效的美容、抗衰、提高身体机能的疗法。
什么是干细胞培养上清液?
首先采取人类脂肪组织→2.进行干细胞分离→3.干细胞增殖培养(干细胞培养上清液)
在干细胞增殖培养过程中,干细胞会分泌出各种细胞因子,所以干细胞培养上清液中富含从干细胞分泌出来的各种细胞因子,这些细胞因子拥有细胞分泌出来的数百种蛋白质,可以调节细胞的增值及分化,使细胞中受损组织及细胞恢复功能受损组织及细胞的恢复及再生是让我们恢复美丽与健康的关键。

干细胞上清液的功能与效果
抗衰老,美容
IGF(胰岛素样生长因子)可促进皮肤再生,有改善皱纹,肌肤弹性再生、增发的效果。
EGF(表皮生长因子)可改善色斑,皮肤暗淡,预防皱纹。
增发,育发
VEGF(血管内皮生长因子)增发,育发
KGF(角质化细胞生长因子)增发,育发
男性功能障碍,组织修复再生,提高免疫力
HGF(干细胞生长因子),PDGF(血小板衍生生长因子),转化生长因子:治疗男性功能障碍,产生抗原抗体反应,有消炎作用及组织修复作用。
预防老年痴呆
NGF(神经生长因子),BDNF(脑源性神经营养因子):促进神经细胞成长,预防老年痴呆。
独特治疗理念的医疗中心
日本银座干细胞诊所,位于东京都中央区银座,该院已获得日本厚生劳动省批准的自体脂肪干细胞临床应用资质。是日本少数获得厚生劳动省认可的两种再生医疗机构,厚生劳动省发布的再生医疗委员会委员。

“再生医学”抗衰与疾病治疗为理念,独特而全面的治疗流程:通过自体脂肪培养,让抗衰老、关节炎、谢顶脱发各类患者都可以享受健康快乐的精彩人生。解决顾客对于容颜和身体上的困扰,更致力于利用安全可靠的再生医疗技术,让患者拥有一个由内而外的健康身体。
多睦健康,提供完善的海外就医服务
从患者进行咨询开始到出国就医再回国,提供完善的就医服务,协助患者完成从病例资料的专业翻译、签证的申请、海外的全程陪同翻译服务,都有着严格的要求和标准。公司有自营海外医疗经验团队,势必成为服务于客户的贴心、专业、全面的海外医疗顾问团队。
分类: 医疗健康
问题描述:
前两种英文字母的是针剂,后两种大概是片剂,不知它们的各自作用是什么,联合使用的相互作用是什么?有没有什么副作用?能不能长期使用?因为是疑癫痫病患者使用,所以比较谨慎。谢谢各位啦!
解析:
神经生长因子(NGF)的基础知识
1. 什么是神经生长因子?
神经生长因子(NGF)是神经营养因子中最早被发现,目前研究最为透彻的,具有神经元营养和促突起生长双重生物学功能的一种神经细胞生长调节因子,它对中枢及周围神经元的发育、分化、生长、再生和功能特性的表达均具有重要的调控作用。NGF包含α、β、γ三个亚单位, 活性区是β亚单位, 由两个118个氨基酸组成的单链通过非共价键结合而成的二聚体,与人体NGF的结构具有高度的同源性,生物效应也无明显的种间特异性。
αNGF亚基 功能尚不清楚(红色)
γ亚基 具有蛋白酶活性(绿 色)
β亚基 具有生物活性的NGF(兰色)
2.研究历程
1953年 意大利科学家Levi-Montalcini发现了NGF。
1960年 美国科学家Cohen提取纯化NGF,证明其生物活性。
1970年 Cohen证明NGF是个复合蛋白。
1984年 NGF的研究重点从周围神经系统拓展到中枢神经系统,乃至非神经系统。
1986年 Montalcini和Cohen因对NGF研究的杰出而荣获诺贝尔生理医学奖。
90年代 国内外多家制药公司和药物研究机构相继开始进行NGF开发研究。
2001年 北京圣日医药科技发展有限公司第一个获得中国SDA颁发NGF新药证书。
2002年 武汉海特生物制药股份有限公司和北京圣日合作开始了NGF产业化进程。
2003年1月6日 世界上第一个神经生长因子——金路捷在武汉海特上市。
3.NGF在机体中的分布?
NGF在人体内主要分布于脑、神经节、虹膜、心脏、脾、胎盘等组织及成纤维细胞、平滑肌、骨骼肌、胶质细胞、雪旺氏细胞等。
4.制备来源
(1)雄性小鼠颌下腺 :与人类NGF有90%同源性
(2)牛精浆
(3)蛇毒
(4)豚鼠前列腺
5.理化特性
(1)7s NGF:分子量接近140kD,沉降系数为7s的复合物.它由α,β,γ三个亚单位和锌离子构成。其生物活性位于β亚单位。β亚单位是2条由118个氨基酸组成的单链,通过非共价键结合而成的二聚体。
(2)2.5s NGF:分子量13~14kD,沉降系数为 2.5s。其结构与β亚基基本相同。故又称β-NGF。
6.受体分类
(1)膜受体:
低亲和力受体:快NGF受体,跨膜糖蛋白,分细胞外部分、跨膜连接区、胞浆部分,具有G蛋白偶联的信号转导功能。
高亲和力受体:慢NGF受体,跨膜糖蛋白,具有酪氨酸蛋白激酶活性。
(2)核受体
7.受体存在的部位
外周神经系统:交感神经元、感觉神经元。
中枢神经系统:胆碱能神经元、肾上腺素能神经元、肽能神经元、脊髓运动神经元、脑干运动神经元、中脑三叉核神经元、视网膜节细胞、小脑部分神经元。
8.神经生长因子的作用机理是什么?
NGF与受体结合,通过受体介导的内吞机制产生内在化,形成由轴膜包绕、含有NGF、并保持其生物活性的小泡,经轴突沿微管逆行转运至胞体,经酪氨酸蛋白激酶、脂酰肌醇钙、内源性环腺苷酸等第二信使体系的转导,启动一系列级联反应,对靶细胞的某些结构或功能蛋白基因表达进行调控而发挥其生物效应。
9.NGF 生物效应
发育期
诱导神经纤维定向生长
*** 胞体和树突的发育
增加神经纤维支配区的密度
促进神经元的分化发育
能使感觉神经,交感神经节数目增加,体积增大,纤维延长
成熟期
NGF和NGF受体的水平和分布都明显减少,主要效应神经对NGF的依赖性也显著降低,只有部分交感神经元仍需依赖NGF存活。
神经损伤后
NGF受体增加,反映在损伤修复过程中对NGF的
需求。
靶区NGF的水平也明显升高。
10. 何谓神经生长因子的神经营养作用?
在胚胎发育的一定时期内,NGF为效应神经元生存所必须。体外实验证实,如果培养液中不加NGF,神经细胞即不能长出轴突,也不能存活。NGF及其受体也广泛分布于中枢神经系统,由海马和脑皮质产生的NGF可通过胆碱能神经逆行运输至前脑基底核,维持胆碱能神经元的存活和功能。在胚胎发育早期,中枢NGF的含量决定胆碱能神经的密度。在无胆碱能神经支配的小脑区和下丘脑,NGF含量也较高,表明除胆碱能神经外,NGF对其他类神经元也有营养作用。
11. 何谓神经生长因子的神经保护作用?
当NGF的效应神经元受到损伤时,例如切断轴突、药物损害,甚至缺血、缺氧等,神经元将发生一系列的病理改变,包括死亡,实验研究证实NGF通过:
(1)抑制毒性氨基酸的释放;
(2)抑制钙离子超载;
(3)抑制超氧自由基的释放
(4)抑制细胞凋亡等机制而明显减轻或防止这些继发性病理损害的发生。
12. 何谓神经生长因子的促神经生长作用?
如上所述,在切断轴突后给予NGF将减少某些神经元的变性与死亡,无疑这将有助于提高轴突再生的可能性。同时它还影响轴突再生开始的时间和参与再生的神经元数目以及再生神经的质量和速度。
13. 神经生长因子在神经系统以外还有哪些作用?
• NGF的非神经系统作用主要表现在影响免疫细胞的活性,进而调节免疫系统功能。神经生长因子不但调节神经系统功能,而且也是一种免疫调节因子。
• NGF可以抑制某些肿瘤的有丝分裂,促其向良性分化。
• NGF促进创口组织的修复反应,促进创口愈合
碱性成纤维细胞生长因子及其对神经组织的生物学效应
puson 汪华侨 庞水发 2004-8-5 19:39:00 中华显微外科杂志 1998年第2期第21卷 综述
关键词: 碱性成纤维细胞生长因子及其对神经组织 生物学效应
早在1940年,Hoftman等在脑和垂体的抽提物中发现一种能够促进成纤维细胞生长的物质。1974年该物被分离纯化,并命名为成纤维细胞生长因子(fibroblast growth factor, FGF)。接着,人们又分离出一种与之高度同源的物质,由于它含有较多的酸性氨基酸碱基,等电点为酸性(5.6),故命名为酸性FGF(aFGF)。先发现的FGF因对酸和热敏感,等电点呈碱性(9.6),则称为碱性FGF(bFGF)。aFGF和bFGF与后发现的int-2、FGF-5、角质细胞生长因子(KGF/FGF-7)、hst-1/kfgf、FGF-6基因表达产物……共9个成员组成FGF家族〔1,2〕。随着bFGF的高度纯化(Bohlen,1984)、测序(Esch,1985,Simpson,1987)和DNA克隆成功(Araham,1986,Gospodarowicz,1984)引入肝素-琼脂糖亲和层析提纯bFGF,可得到纯度达90%以上,由此bFGF研究进入了新阶段。现今资料表明,bFGF的生物学作用极其广泛,它在血管形成、促进创伤愈合与组织修复、促进组织再生和神经组织生长发育过程中起着十分重要的作用。现就bFGF的一般性质、分布、对神经组织的作用和机制以及临床应用前景等作一综述,以促进bFGF的开发和临床应用。
一、bFGF的一般性质
bFGF是含155个氨基酸的促有丝分裂的阳离子多肽,其氨基酸序的55%和aFGF相同,分子量为16~18.5 KD。bFGF分子结构中有4个半胱氨基酸,以此形成分子的三维空间结构。由丝氨酸取代半胱氨酸的重组bFGF的生物学活性不变,而单链多肽则易于在大肠杆菌中表达〔3〕。
bFGF基因位于人的第5对染色体上,为单拷贝基因,呈不连续状态,其功能区被两个内含子隔开分为三个外显子区域。bFGF基因长度大于38 kb,第一个内含子位于第60、61位密码间,第二个内含子位于第94、95位密码子之间。bFGFmRNA有4.6 kb和2.2 kb两种形式,有bFGFmRNA反转录的cDNA序列已清楚,在bFGFcDNA可读框架中可见到一个常见的AUG起始密码、UGA终止密码。在AUG前面有起始密码CUG存在,它可启动氨基末端的延长。bFGF羧基末端较氨
基末端稳定,如果氨基末端截取少于25个氨基酸时并不影响其生物学活性。bFGF活性比aFGF大30~100倍〔4〕。bFGF在生物进化上具有很强的保守性,各种动物的FGF都有很高的同源性。人和牛bFGF氨基酸顺序同源性达98.7%。
bFGF有很强的肝素亲和力,在其114~123位氨基酸为高亲和力区,其它部位则有低亲和力区。抗bFGF与受体结合的单克隆抗体对其与肝素结合力无影响,取消有羟基端42位氨基酸,肝素亲和力即消失,而且可丧失部分生物学活性。bFGF基因中未发现信号肽顺序,用bFGF的cDNA转染3 T3细胞,能观察到bFGF对单个细胞的趋化作用,且用抗bFGF的mcAb可中和这一活性,说明bFGF可经自分泌方式释放〔5〕。
二、bFGF的分布
主要分布于垂体、脑和神经组织及视网膜、肾上腺、胎盘等〔17〕,尤以垂体含量最高,能纯化大量的bFGF(0.5 mg/kg),其它组织含量很少,约为其1/10~1/50。bFGF不存在或以极低浓度存在于血清和体液中。
bFGF作为细胞分裂原,主要作用在起源于中胚层和神经外胚层的骨骼肌细胞、成纤维细胞和骨细胞等,其受体也相应的分布于上述细胞表面。FGF存在两类受体:一类是亲和力受体,属跨膜性酪氨酸蛋白激酶类受体;另一类是低亲和力受体,即肝素样受体,为硫酸乙酰肝素蛋白多糖类物质〔6〕。它们是一条单链多肽,约110~150 KD,受体数目约2×103~8×104/细胞,受体对bFGF的亲和力KD=18~80 pm。受体至少有4种形式,由细胞外区、跨膜区、胞浆区的近膜区和酪氨酸激酶区组成,由于每种FGF受体均能和FGF家族每个成员结合,而不同FGF受体的表达存在着组织细胞特异性。bFGF与受体亲和力显著大于aFGF。
三、bFGF的作用机制
bFGF通过与靶细胞上的受体结合而发生作用,因此细胞内合成的bFGF需分泌至细胞外才能发挥生物学作用。但bFGF的mRNA翻译产物缺少引导它们向细胞外分泌的信号序列,其分泌途径与经典途径不同,除了可能是细胞受损或死亡后释出〔7〕,还有自分泌和旁分泌起作用〔8〕。
bFGF与高亲和力受体结合时需低亲和力受体的参与,提示低亲和力受体的结合使高亲和力受体结合更容易、更牢固〔9〕。bFGF与受体结合后可能通过以下途径将信号传至胞核:(1)激活腺苷酸环化酶与鸟苷酸环化酶,磷脂酶C(PLC-rl)磷酸化,又使磷脂酰肌醇-4,5二磷酸(PIP2)分解为甘油二酯(DG)和三磷酸肌醇(IP3),导致蛋白激酶C激活和Ca2+内流;(2)与受体结合后定位于细胞核,影响RNA聚合酶Ⅰ,加强 *** 白体基因的转录,以加速细胞由G0→G1和→G1→S期的转换, *** 细胞的DNA合成增强,促进细胞的分裂与增殖〔9〕;(3)bFGF与FGFR复合物的内部化。
四、bFGF的生物学功能
bFGF的生物学效应分体内和体外两大部分。体外作用十分强烈,成纤维细胞、骨细胞、软骨细胞、血管内皮细胞、肾上腺皮质和髓质细胞、神经元和神经胶质细胞等具有很强的促细胞分裂增殖活性〔10〕。体外细胞培养中能在低浓度(1 mg·ml-1)发挥其作用。bFGF是重要的促有丝分裂因子,也是形态发生和分化的诱导因子〔11〕。其主要生物学作用有:(1)作为血管生长因子;(2)促进创伤愈合与组织修复;(3)促进组织再生;(4)参与神经再生等。
五、bFGF对神经组织的生物学效应
1.bFGF在神经组织的表达
从多种神经外胚层和中胚层起源的组织(如大脑皮质、下丘脑、垂体和视网膜等)可提取、纯化出bFGF。采用免疫组织化学方法测出神经元胞体、轴突与树突近端bFGF浓度为40~120 pm*g-1〔12〕。在星形胶质细胞和海马神经元、腰段脊髓神经元、神经胶质细胞及坐骨神经的雪旺氏细胞、郎飞氏结等也发现有bFGF的分布〔13,14〕。在鹌鹑的胚胎期,发现神经管和神经嵴有bFGF表达,后期在脊索和脊索神经节根表达。
中枢神经损伤后,有关bFGF表达情况的研究较多。Finklestine等(1988)发现脑损伤后bFGF明显增加,尤其病灶周围星形胶质细胞最为显著;Salley(1991)报道大脑损伤后3天,患侧皮层、室管膜和海马神经元中bFGF增加;Christine等(1994)诱发成年大鼠癫痫发作时,前脑神经元和胶质中bFGFmRNA显著增多。但在周围神经有关神经损伤后bFGFmRNA的表达,尚未见文献报道。
2.bFGF作为神经营养因子
(1)是神经胶质细胞和雪旺氏细胞的促有丝分裂原。bFGF有 *** 神经胶质细胞的非有丝分裂活性,如促进星形胶质细胞的迁移和纤溶酶活剂的释放;调节胶质细胞纤维酸性蛋白(GFAP)的表达及谷氨酸和S-100蛋白的合成;改变星形胶质细胞的典型的细胞进程和细胞膜结构;促进星形胶质细胞的增殖并形成纤维状外形;也可促进少突胶质细胞的增殖,并增加其髓磷脂相关蛋白和类脂的含量。
(2)对体外培养神经元的作用
bFGF能延长培养液中多种中枢和外周神经元的存活, *** 胆碱乙酰化酶的合成以及突起的生长。Aoyagi等〔15〕报道在培养的胎鼠海马神经元中加入bFGF,可使神经元成活时间增加和其轴突延长。在培养的胎鼠海马神经元中加入bFGF10~30pg·ml-1,使原只能存活5~7天的神经元生命延长14天,数目增加4倍;当浓度增至200~500pg·ml-1时,可使原只30μm的突起延长至100μm。Patner等(1988)在培养的雪旺氏细胞中加入bFGF后,5%~10%的细胞进入分裂期。bFGF对培养中的胚鼠脑的额区、顶区、纹状体、丘脑的胆碱能神经元和多巴胺能、γ-氨基丁酸能神经元,大鼠小脑皮质神经元、交感节细胞、鸡胚脊髓前角神经元等都有营养和促进作用。
(3)体内神经营养作用
当bFGF用于损伤的大脑时,能促使海马神经元存活,而无bFGF时海马神经死亡。在外周神经系统,当bFGF加入紧靠坐骨神经切断处,能够促进神经的髓鞘化,防止背根神经节神经元的死亡〔16〕。Seivers(1987)、Gospodarowicz(1990)等也证实bFGF可使切断视神经后的视网膜后的视网膜节细胞成活。将bFGF注入大鼠脑中,也可保持切断轴突的大脑皮层的胆碱能神经元的存活(Anderson,1988)。Ferrari等(1989)经体内实验证实,bFGF能提高中脑腹侧多巴胺能神经元移植物的成活。
(4)对周围神经再生的促进作用
在周围神经损伤修复的研究中,有资料表明,bFGF有明显的促进外周神经纤维再生的作用是比较肯定的,也已在体的神经“套管”模型实验中得到证实〔16〕。自Lundb(1982)建立神经再生模型后,有关加入某些因子对神经再生影响的研究很多。Cuevas〔17〕等给切断坐骨神经灌注bFGF,可提高神经的再生率。Laquerriere等〔18〕在桥接大鼠7mm长坐骨神经缺损中使用bFGF,4周后发现神经再生成功。Koshinaga等〔19〕研究了脊髓损伤后bFGF、aFGF的表达情况后认为,aFGF、bFGF参与脊髓损伤的修复过程。有研究表明bFGF的促神经再生作用可与NGF家族、睫状节神经营养因子(CNTF)、胰岛素样生长因子(IGFs)等的神经营养活性相互协同〔2〕。
(5)促进神经前体细胞分化
bFGF有对神经前体细胞的增殖分化作用。Gen *** urgeror(1987)发现培养的大鼠神经元加入bFGF后,出现胆碱能成份分裂并增殖。Dicico-Bloom(1990)观察到成神经细胞的分裂受bFGF的调节,分裂过程中出现轴突突起生长出生长锥、神经递质合成、递质小泡的转运等神经元特性。此外,bFGF还可通过它的促血管生成作用来影响中枢神经和周围神经系统的发育。
六、存在的问题和临床应用前景
bFGF是一种促细胞分裂的肝素结合蛋白,可诱导多种细胞的增殖与分化,对神经系统有重要作用。在不同种间bFGF结构的高度守恒性,提示它在个体发育中起着原始的促进作用。鉴于以往bFGF的研究多集中于中枢神经系统,在周围神经损伤后,bFGFmRNA的表达情况、bFGF受体表达的细胞、bFGF如何与受体识别以及结合后的变化?bFGF促周围神经再生的机制,生理状态下存在多种生长因子,它们的相互协调作用以及调节关系怎样?等,这些问题的阐明将为临床提高周围神经损伤后的修复效果提供理论依据。此外,由于bFGF易被酶分解,其在体内作用的发挥需持续长时间与靶细胞受体结合,如研究一种既能让bFGF稳定不受蛋白酶降解,又能使bFGF持久缓慢释放的载体,则解决了bFGF临床的一大难题。虽然bFGF在体内含量甚微,但分布广泛,生理功能复杂多样。bFGF生物活性的多效性以及神经营养的广谱性,为其从基础走向临床提供了保证,bFGF对于神经损伤再生的研究,是对于神经损伤治疗领域的一个新的探索和拓宽,目前在动物实验上已现成效。国内第二代基因重组h-bFGF也已经问世〔4〕,它的出现,展示着bFGF临床应用的光明前景。
参考文献
1 左藤靖史.血管新生の诱导因子と阻害因子.最新医学,1995,50(6):25-30.
2 周廷仲主编.多肽生长因子基础与临床.第1版.北京:中国科学技术出版社.1992,209-230.
3 Arakawa T. Characterization of a cystein-efree
*** og of rebinant human basic fibroblast growth factor. Biochem Biophys Res Commun,1989,161:335-341.
4 林剑.碱性成纤维细胞生长因子.暨南大学学报,1993,14:99-103.
5 Mignatti P. Basic fibroblast growth factor released by single isolated cells stimulated their migration in an autocrine manner. Proc Natl Acad Sci (USA),1991,88:11007-11011.
6 Eckenstein FP. Fibroblast growth factors in the
nervous system. J Neurobio,1994,25:1467-1471.
7 Muthukrishnan L, Warder E, Mchelt PL. Basic fibroblast growth factor is efficiently released from a cytolsolic storage site through pla *** a membrane disruptions of endothelial cells. J Cell Physiol,1991,148:1-16.
8 Mignatti P, Morimoto T. FGF: a protein devoid of secretory signal seguence, is release by cells via a pathway independant of the endopla *** ic reticulum-golgi plex. J Cell Physiol,1992,151:81-93.
9 Jaye M, Schlessinger J, Craig A, et al. FGFR tyrosine kinases molecular *** ysis and signal transduction. Biochem Biochem Acta,1992,1135:185-199.
10 Coltrini D. Biochemical bases of the interaction of human basic fibroblast growth factor with glycosamine-glycans. Eur J Biochem,1993,214:51-58.
11 Doniach T. Basic FGF as an inducer of anteroposterior neural pattern. Cell,1995,83:1067-1072.
12 Pettman B, Labourdette G, Weibel M, et al. The brain fibroblast growth factor is Localized in neurons. Neurosci Lett,1986,68:175-181.
13 Eckenstein FP, Kuzis K, Nishi R. Cellular distribution subcellular localization and possible functions of bFGF and aFGF. Biochem Pharmacol,1994,47:103-111.
14 Hassen SM, Kerkhoff H, Troost D. bFGF immunoreactivity in the peripheral motor system of the rat. Acta Neuropathol Berl,1994,87:405-411.
15 Aoyagi A, Nishikawa K, Sait OH. Characterization of bFGF-mediated acceleration of axonal bronching in cultured rat hippocampal neurons. Brain Res.1994,661:117-123.
16 Denielsen N. Fibroblast growth factor effects on peripheral nerve regeneration in a silicone chamber modil. J Neurosci Res,1988,20:320-330.
17 Baird A, Walicke PA. FGFs. Med Bull,1989,45:438-454.
18 Laquerriere A, Peulve P, Jin O. Effects of bFGF and alpha-melanocytic stimulating hormone on nerve regeneration through a collagen channel. Microsurg,1994,15:203-211.
19 Koshinage M, Sanon HR, Whittemore SR. Altered acidic and basic fibroblast growth factor expression following spinal cord injury. Exp Neurol,1993,120:32-41.
扶济复®
外用重组人碱性成纤维细胞生长因子 ,rh-bFGF
国家一类新药
●全面促进组织创伤的修复;
●生产工艺先进,稳定性好;
●国家行政保护,12年独家生产。
【适应症】
促进创面愈合,可用于烧伤创面(包括浅II度、深II度、肉芽创面)、慢性创面(包括慢性肉芽好、溃疡和褥疮等)和新鲜创面(包括外伤、手术伤等)。
【规 格】 20000AU/支
扶济复
外用重组人碱性成纤维细胞生长因子(rh-bFGF) 20000 AU (35000IU)批发价 103.57 零售价 116.00